

Las paredes celulares son la principal reserva de carbono en el planeta. Este carbono, en forma de estructura muy diversa, tiene un potencial de aplicación muy grande, pero el problema es obtenerlo en sus diferentes estructuras químicas para su uso. La deconstrucción de las paredes celulares de las plantas, usando enzimas capaces de degradar específicamente cada componente, tienen un potencial de uso muy grande para un sinnúmero de aplicaciones industriales, médicas, nutricionales, nuevos materiales, etc.
Cuando éramos estudiantes, llevábamos cursos de botánica en donde se incluían las bacterias, líquenes, hongos y otros microrganismos. No es que los creyeran plantas, el asunto es que no se sabía cómo ubicarlos, y como estos grupos tienen pared celular como una estructura que envuelve a las células, entonces era común ubicarlos con las plantas y listo, una clase de botánica criptogámica. Se parecían a las plantas, pero carecían de flores.
Con el tiempo se retiraron de este grupo, puesto que, aunque tengan pared celular posee una composición, estructura y origen distinto. No es nuestro propósito discutir estas clasificaciones, sino llamar la atención de cómo una estructura muy evidente puede suponer confusiones. La pared celular vegetal, ciertamente es una estructura muy notoria en las plantas y, junto a los grupos indicados, las separa de los animales.

Por pared celular (PC en adelante) entendemos una estructura externa que rodea la célula. Está constituida por diversos tipos de moléculas sintetizadas por la misma célula a manera de una matriz que se exporta al exterior y se consolida como una especie de bolsa. En ocasiones llega a tener material incorporado externo, básicamente minerales que contribuyen con sus propiedades. Esta estructura limita el crecimiento celular y, en el caso de las plantas, cuando se vuelve muy rígida, la célula muere y solo queda el «fantasma» de las paredes a manera de tubería microscópica.
La PC se ha visto, y de hecho se sigue enseñando así, como un material rígido, inerte y que básicamente tiene funciones mecánicas y de soporte. Esta es una noción equivocada, ya que en realidad estamos frente a una estructura muy dinámica, la cual cambia a lo largo de la vida de la planta, del tejido u órgano que se trate y de acuerdo con las condiciones ambientales.
Imaginemos un girasol. Esa planta debe su nombre a un comportamiento muy curioso. Su flor se mueve cual antena parabólica siguiendo la posición del sol. Estamos tan acostumbrados a ese comportamiento —en esa y cientos de otras plantas—, que no meditamos la cantidad de procesos que deben ocurrir para que ese movimiento sea posible, y es que una planta no tiene un sistema muscular que articule el movimiento… Continuando con el ejemplo del girasol, pensemos que su pedúnculo, la ramilla que sostiene a la flor, está formada por miles y miles de células contenidas en una estructura muy fuerte y rígida que debería impedir el movimiento, pero es flexible y eso le permite sortear el viento, el roce de otros organismos, incluso la tensión provocada por obstáculos.
¿Entonces, cómo se mueve?
El secreto está en los cambios de la tasa de degradación y síntesis de la PC. La luz activa fotorreceptores que, al percibirla, inducen la síntesis y exportación de enzimas involucradas en la fabricación de la PC, pero al lado opuesto de donde viene esta luz. La síntesis va acompañada de un arreglo nuevo del citoesqueleto que trasportará estas enzimas y monómeros de azúcares (ladrillos) que formarán las fibras de la PC en vesículas desde el citoplasma. Adicionalmente, se produce el bombeo de iones H+ en ese lado opuesto a la luz, lo cual acidificará el medio, provocando la activación de las enzimas que degradan la PC y que previamente fueron exportadas al espacio intercelular, tales como las enzimas que degradan celulosa o pectina. Estas enzimas realizarán cortes a las fibras permitiendo que se inserten nuevos ladrillos, alargando la PC de ese lado y aumentando de manera asimétrica su longitud, lo cual inclinará la rama hacia donde viene la luz. El fenómeno ocurrirá del otro lado cuando la posición del sol cambie. Sofisticado, ¿no lo creen? Seguro han notado que las plantas crecen más y se alargan mucho en la búsqueda de la luz.
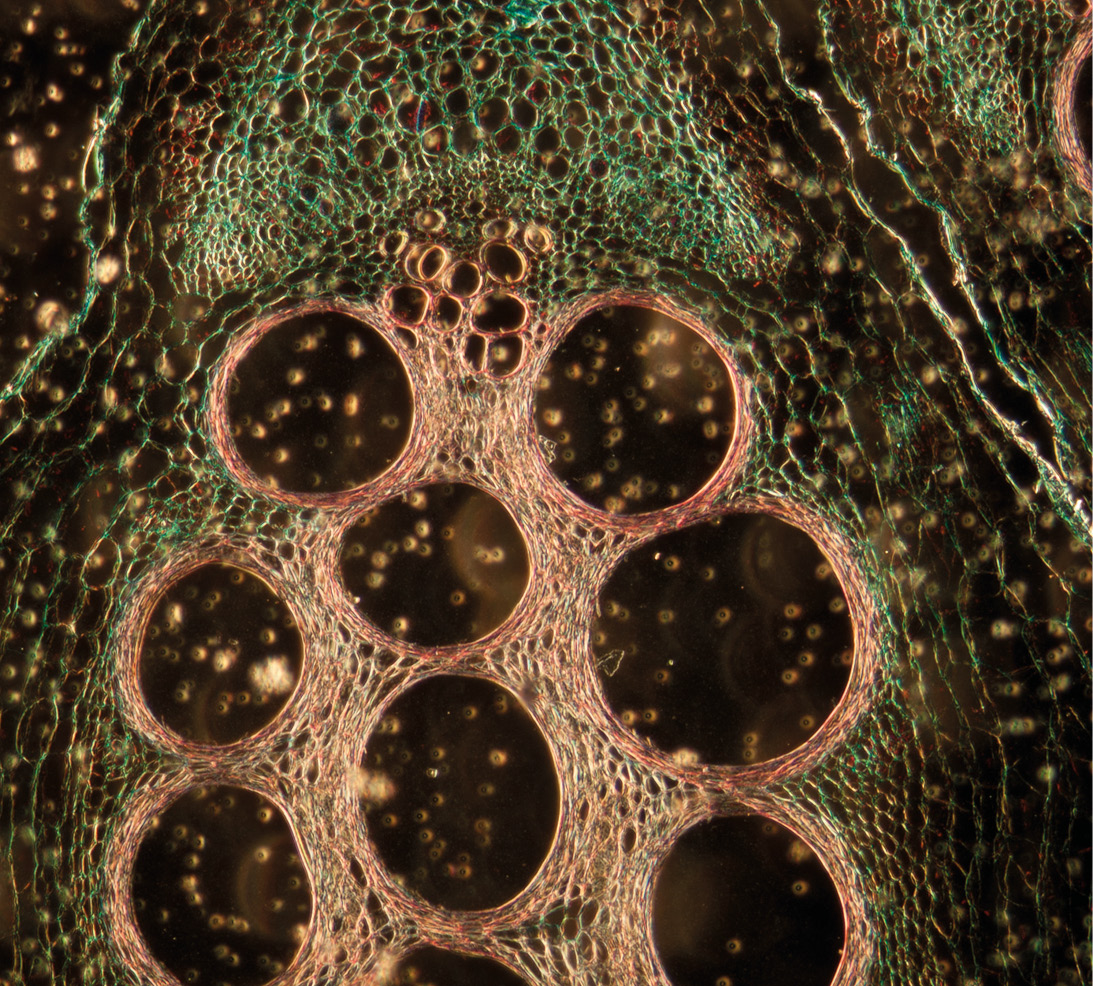
Composición y estructura de las paredes celulares vegetales
Si bien la PC de todos los organismos que la presentan tiene algunas funciones similares, su composición, estructura y síntesis es muy diferente: los biólogos decimos de este fenómeno que son estructuras «análogas», pero no «homólogas».
Las PC vegetales se forman mayoritariamente de carbohidratos, es decir, cadenas de azúcares (80 %). Además de esta porción, acumulan lignina, un polímero de alcoholes aromáticos y también se acumulan proteínas. Además, pueden agregarse, dependiendo de la planta, calcio (Ca+2), fierro (Fe+2) y otros iones divalentes. Esta acumulación de metales contribuye a la dureza de la pared celular en algunas plantas. Seguro han escuchado hablar del «palofierro» (Olneya tesota), un árbol de madera sumamente dura que se oxida como un clavo y, de hecho, desarrolla el color de metal oxidado y es utilizado por los pueblos Seris y Yaquis del noroeste de México para realizar artesanías. Resulta que esta madera está hecha de PC endurecidas.
Pero las PC no son una acumulación de carbohidratos y otros componentes tal como en una masa. En realidad, poseen una estructura compleja y diversa. De entrada, podemos clasificar al material del que están hechas en fibras y matriz de la pared. Las fibras representan el 80 % del material y básicamente son polisacáridos como la celulosa (30 %), las hemicelulosas (30 %), las pectinas y otras glucanas (compuestos poliméricos, generalmente de pequeño tamaño de azúcares como glucosa, manosa, galactosa, etc.). Los porcentajes son relativos puesto que varían con la planta, la edad, el tejido, etc. Las hojas verdes, por ejemplo, presentan mayores porcentajes de hemicelulosa (lechugas, alfalfa, etc.), mientras que hojas y tallos fibrosos como el maíz y la caña de azúcar, tienen mayor contenido de celulosa junto a lignina. Por su parte, la matriz está formada por hemicelulosas, pectinas, lignina, otros polifenoles y la fracción proteica.
Vamos a revisar la estructura y para ello haremos uso de una analogía. Seguro todos los lectores conocen o han visto el concreto armado, también llamado hormigón reforzado, el cual resulta de la combinación de un material cementante (mezcla de agua, arena, cemento, cal que al mezclarse y secarse adquiere mucha dureza) con la presencia de barras de acero. Al mezclarse forman elementos estructurales como vigas, planchas, muros, columnas, etc. Las barras de acero (varillas) están a su vez conectadas con material que mantiene su acomodo al recibir el concreto.
Las PC vegetales tienen una estructura análoga. Existen varillas representadas por la celulosa, incluidas en una matriz de diversos polisacáridos y otros materiales que en nuestra analogía sería el concreto. La celulosa es un polímero de glucosa formado por hasta 20,000 unidades de glucosa que constituyen una cadena. Luego, una cadena se une a otra por atracción electrostática y junto con 200 de estas cadenas forman una microfibrilla, que es la unidad básica funcional de la celulosa. Hay millones de estas microfibrillas formando una red extensa que se entrecruza de manera tridimensional, tal como ocurre con la estructura de acero del concreto armado.
Las microfibrillas están «amarradas» por otros polisacáridos, básicamente las llamadas hemicelulosas formadas por xilosa, ramnosa, galactosa, etc. Que se unen formando fibras más cortas (xilanas, xiloglucanas, ramnoxiloglucanas y mucha más diversidad) y que se encuentran altamente ramificadas. Las hemicelulosas mantienen unidas a las fibras de celulosa y además la protegen del daño por factores externos, tales como fitopatógenos, además de impedir la disposición al azar de la celulosa.
En las zonas de contacto célula-célula se acumula mayoritariamente otro material polisacárido, las pectinas. Esta zona se conoce como lámina media. Una diferencia notable de las pectinas y las hemicelulosas es que estos últimos se forman de azúcares sin carga, mientras que las pectinas están formadas por un esqueleto de azúcares cargados, residuos del ácido galacturónico (un azúcar ácido) en su esqueleto principal, alternándose con la ramnosa y ramificaciones de esta. Esto es importante porque provoca que las pectinas estén fuertemente hidratadas y que, además, puedan atraer iones divalentes positivamente cargados, tales como el Ca+2 que ya mencionamos. Este compuesto forma una estructura semirrígida, porosa y con las propiedades de un gel, muy importante en la matriz de las PC.
La flexibilidad, unida a una fuerte resistencia, también se ve reforzada por la presencia de proteínas. Podemos encontrar —si bien sus cantidades son muy variables—, dos grupos de proteínas: las llamadas extensinas y un grupo llamado proteínas ricas en hidroxiprolina; por cierto, este tipo es muy similar al colágeno de los animales. Las proteínas proporcionan resistencia a la tensión y a la digestión por herbívoros, de manera que tienden a acumularse en la pared como una respuesta al daño mecánico y la herbivoría. Dependiendo del tipo de tejido y la edad, en la matriz también se pueden incorporar glucanas, una gran diversidad de polímeros y oligómeros de glucosa, manosa, ramnosa, entre otros, que refuerzan el material cementante.
El otro gran componente estructural de las PC vegetales es la lignina. Este es un polímero no lineal formado por alcoholes aromáticos como el cumarílico, coniferílico y sinapílico. Estos compuestos, digamos de tamaño pequeño, se van uniendo para formar una macromolécula muy extensa, insoluble y de muy difícil digestión, que además contribuye a darle resistencia y dureza a la PC.
Podemos distinguir dos estadios de desarrollo de las PC, dependiendo de la edad de la planta. Cuando los vegetales están en una fase de crecimiento rápido, con tejidos verdes y en general células vivas, la pared está en un estado de crecimiento y reconfiguración constante llamado paredes celulares primarias. En este estado hay poca lignina y esta, junto a otros polifenoles, solo se acumula como respuesta al daño o al ataque de patógenos y herbívoros. Sin embargo, a medida que la planta deja de crecer —fuera de las células en división (meristemos apicales) —, muchos tejidos comienzan a acumular lignina y a sustituir otros azúcares por esta, formando complejos matriciales con las fibras de celulosa conocidos como lignocelulosa. La PC se vuelve muy rígida, incluso en una etapa llamada de paredes celulares secundarias. En esta etapa generalmente podemos identificar a las células como los «fantasmas» de células preexistentes y las paredes forman «tuberías» por donde circula agua. Este tipo de material lo encontraremos en tejidos muy fibrosos y resistentes como los rastrojos, la paja, el xilema y desde luego, la madera.
Hemos realizado una descripción muy general de la composición y estructura de las PC vegetales, debido a que es una estructura dinámica que cambia, como ya lo dijimos, con la edad de la planta, el tejido y como respuesta al estrés ambiental, reforzándose con material adicional, degradando ese material para proporcionar azúcares y acumular pigmentos, como en el proceso de maduración de los frutos. Podemos calcular el porcentaje de polisacáridos y otros componentes en diferentes plantas y tejidos, pero estos cambian mucho y eso hace a las PC complejas de estudiar, aunque fascinantes.
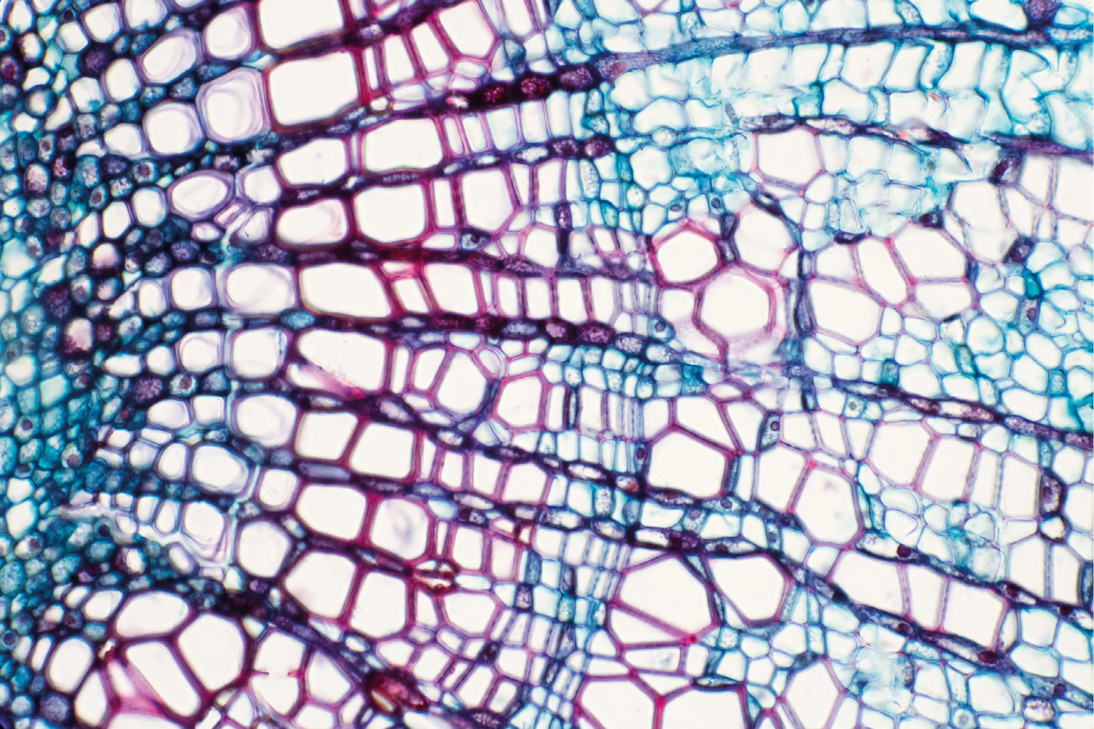
Funciones de las paredes celulares vegetales
Una estructura cuya función es únicamente proporcionar soporte y resistencia no necesita ser tan dinámica. Durante mucho tiempo esta fue la concepción dominante; sin embargo, esto comenzó a cambiar con la descripción del involucramiento de la pared celular en procesos tales como la defensa contra patógenos, la resistencia a la herbivoría, la floración, la maduración de frutos, entre otras funciones.
Cierto, es una estructura muy resistente que, además, rodea las células, las protege de la presión de los líquidos contenidos en la célula, regula el paso de moléculas y le da forma a la célula.
Sabemos en la actualidad que un mecanismo para penetrar a las células de las plantas y alimentarse de ellas de parte de los patógenos, es el uso de enzimas que rompen los polisacáridos de la PC. Esto ablanda el muro de protección, permitiendo penetrar a las células y tejidos, y proporciona alimento. En el proceso se liberan pequeñas moléculas de los polímeros que actúan como señales del daño y estos son reconocidos por receptores en las membranas que articulan respuestas de defensa, como la respuesta celular hipersensible (muerte celular programada), la síntesis y acumulación de antibióticos, lignina y polifenoles, la acumulación de proteínas relacionadas con la resistencia y la generación de otras señales que viajan a largas distancias, advirtiendo del peligro potencial y preparando a la planta y a las plantas vecinas para la defensa.
Estos fragmentos u oligosacáridos también están involucrados en la apoptosis o muerte celular programada, involucrada en la defensa, caída de frutos, flores, la misma floración y, junto con otras fitohormonas, regulan una gran diversidad de respuestas del desarrollo de la planta.
Potencial biotecnológico de las paredes celulares vegetales
Las PC son el principal reservorio de carbono en la tierra. De hecho, son el origen del petróleo y el carbón mineral, los diamantes, el grafito y por supuesto, el proceso organizador de la vida en el planeta, la fotosíntesis que consiste en la captura de carbono de la atmósfera. Nos alimentamos, todos los seres vivos, para obtener carbono y realmente somos un planeta de carbono, la vida es la química del carbono.
Imaginemos cuanto carbono hay almacenado en la PC con un cálculo muy sencillo. Ya dijimos que la celulosa, más o menos el 30 % de los carbohidratos de la pared, se forma de microfibrillas. Cada fibrilla está formada por 200 cadenas de glucosa, cada glucosa tiene seis carbonos, entonces en una sola cadena hay aproximadamente 120,000 átomos de carbono, como cada microfibrilla tiene 200 de estas cadenas, entonces una sola microfibrilla posee 24,000,000 de átomos de carbono. En una pared, de una sola célula, hay miles de millones de microfibrillas, es decir, cantidades gigantescas de carbono almacenado en cada planta, por pequeña que sea, y a eso hay que sumarle los otros polisacáridos y componentes de esta estructura sorprendente. Hagan cálculos…
Una estructura tan compleja como la PC, ofrece infinidad de usos. De entrada, la madera, el papel, las fibras vegetales (lino, algodón, cáñamo, etc.), solventes, resinas, texturizantes de alimentos, entre otros, conocidos desde hace siglos.
Pero el entendimiento de la composición y la dinámica de las PC, ofrecen usos potenciales muy amplios. Hablábamos de la PC como fuente de señales para controlar aspectos del desarrollo, como la defensa, crecimiento y floración. También es una fuente de fibras alimenticias, los llamados carbohidratos complejos: las hemicelulosas, las pectinas y otros polisacáridos de las PC son macromoléculas que nosotros no podemos digerir y de muy difícil digestión por los microrganismos del microbiota intestinal. Eso tiene varios impactos positivos: contribuye a la digestión de otros alimentos; dificulta la absorción de azúcares simples, disminuyendo el índice glucémico de muchos alimentos; el incremento de la sensación de saciedad y como en nuestro tracto digestivo tenemos bacterias que sí pueden digerir estos carbohidratos complejos, pero lo hacen muy lentamente, los azúcares necesarios para el metabolismo se liberan lentamente, además, estos son distintos a la glucosa, lo cual activa otras rutas metabólicas contribuyendo a la nutrición, pero evitando la hipoglucemia y la producción de insulina.
Las fibras tienen otros usos potenciales, desde la elaboración de nuevos materiales, en particular los llamados nanomateriales, hasta la elaboración de «madera sintética» y todos los usos que imaginemos, con el consecuente efecto positivo al fenómeno de deforestación, ya que la idea es incorporar fibras de plantas no maderables, desechos de la agricultura, la agroindustria, material de malezas y un largo etcétera.
También podemos obtener de los carbohidratos complejos, edulcorantes indigeribles o poco digeribles como la xilosa, xilitol, etc., que conserven lo dulce, pero no impacten en la concentración neta de glucosa, compuestos inmunorreguladores, gomas, aceites, biocombustibles, etc.
El problema radica en que las PC vegetales constituyen un material sumamente recalcitrante a su purificación. De hecho, la preparación de papel que está hecho básicamente de celulosa, implica el uso de solventes, quelantes, ácidos, además de calor y mucho trabajo mecánico para liberar las fibras puras de celulosa, con la consecuente liberación de muchos desechos, como los derivados de la lignina, que contaminan mucho y llegan a ser muy tóxicos.
De manera que, debemos buscar formas alternativas y aquí la naturaleza misma marca el camino. Existen muchos organismos que sí pueden digerir la PC y lo hacen con procesos enzimáticos muy específicos. Entre estos organismos hay bacterias celulolíticas (así se les llama, pero pueden digerir otros polisacáridos, además de la celulosa), hongos, tanto fitopatógenos, como saprófitos (que se alimentan de plantas muertas), nemátodos, protozoarios y los insectos xilófagos (que se alimentan de madera).
En nuestro grupo de investigación, nos interesan particularmente los hongos ya que presentan una «batería» completa de enzimas que degradan cada componente de la pared celular de forma muy específica.

Deconstruyendo la pared celular vegetal
Existe una cantidad muy amplia de hongos cuyo estilo de vida depende de su habilidad para digerir y romper las PC. Los hongos fitopatógenos requieren de las llamadas enzimas activas sobre carbohidratos (llamadas CAZymas) para ablandar las paredes hasta romperlas. Para ello, usan la fuerza mecánica producida por incrementos en la presión de turgencia de las hifas (cuerpo de los hongos) por acumulación de glicerol y melanina. Esta presión impacta directamente sobre las PC, pero va acompañada de la secreción de CAZymas que van cortando las fibras de las PC, además son capaces de degradar la lignina y otros componentes en un símil a un taladro que va perforando. Los hongos saprófitos obtienen su energía de la materia vegetal muerta degradando las PC para obtener carbono. Ambos estilos de vida requieren una gran cantidad de CAZymas.
En nuestro grupo usamos de modelo a un hongo patógeno del frijol llamado Colletotrichum lindemuthianum, responsable de la principal enfermedad de este cultivo, la antracnosis. Lo interesante de este hongo es que hemos identificado en la misma especie diferentes aislados o variantes (se les llama patotipos), según su capacidad de infectar al frijol. Y contamos con una serie de patotipos, desde los muy patógenos, hasta variantes incapaces de infectar por sí mismas y que presentan estilos de vida más cercanos a los saprófitos. Imagine contar con un gradiente de estilos de vida en una misma especie.
Con este modelo, nos hemos propuesto identificar y caracterizar todo el proceso de degradación de las PC vegetales, caracterizar y purificar las CAZymas, y conocer el orden preciso en que este organismo va desensamblando las paredes de su huésped, el frijol, para infectarlo. Además, nos interesa comprender la evolución del estilo de vida para comprender no solo cómo se han desarrollado de saprofitos a fitopatógenos o viceversa, sino cómo diferentes patotipos actúan concertadamente para digerir las PC. Este entendimiento, permitirá diseñar sistemas de digestión controlada de las PC, permitiendo purificar cada componente de interés de manera diferenciada y con procesos más amables con el ambiente.
Al momento, hemos purificado, clonado los genes y estudiado su evolución de CAZymas como celulasas, pectinasas, pectinliasas, pectatoliasas, xilanasas, xilosidasas, arabinoxilosidasas y arabinofuranosidasas, entre otras, que permiten ir pensando en sistemas de digestión de diferentes materiales, además de realizar estudios comparativos, sin salir de la misma especie, de la eficiencia catalítica de enzimas provenientes de hongos con estilo de vida diferente, incluso contrastante. Esto es importante, porque en la actualidad la fuente de CAZymas disponible comercialmente son básicamente tres hongos Trichoderma, Aspergillus y Penicillium y se desconoce por completo si son las más adecuadas para todo tipo de materiales vegetales. Nuestra hipótesis, derivada de los resultados experimentales, es que no, que no existen «enzimas universales» eficaces contra cualquier material y que es necesario diseñar nuevos protocolos basados en el conocimiento, evolución y estilo de vida de la fuente de CAZymas respecto a la especie vegetal, cuyo material se quiere procesar.
Finalmente, estos estudios permiten, mediante tecnología del ADN recombinante, diseñar sistemas de producción de enzimas que tienen múltiples aplicaciones industriales, desde el biopulpeo (papel), producción de biocombustibles de tercera generación, aclaramiento de jugos y vino, producción de telas, producción de carbohidratos complejos, nutracéuticos y muchas otras aplicaciones.
El estudio de las enzimas que degradan lignina y derivados y que nuestro hongo modelo tienen en abundancia, también es otra fuente de aplicaciones biotecnológicas que no hemos abordado aún.

María Guadalupe Zavala Páramo. Profesora e Investigadora Titular «C» de Tiempo Completo del Centro Multidisciplinario de Estudios en Biotecnología de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Michoacana de San Nicolás de Hidalgo.
Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.
Horacio Cano Camacho, Profesor Investigador del Centro Multidisciplinario de Estudios en Biotecnología y Jefe del Departamento de Comunicación de la Ciencia de la Universidad Michoacana de San Nicolás de Hidalgo.
Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.