

Lamarck (Jean-Baptiste Pierre Antoine de Monet Chevalier de Lamarck) fue un naturalista francés, que formuló la primera teoría de la evolución biológica, la que en forma resumida podríamos enunciar de la siguiente manera:
-el uso frecuente y sostenido de un órgano, lo fortifica y hace que se desarrolle, por el contrario, sí un órgano no es usado tiende a desaparecer, y estas modificaciones sufridas a lo largo de la vida del individuo son heredadas a generaciones siguientes-
A esta idea también se le conoce como la teoría del “uso y desuso”, cuyo ejemplo clásico es la explicación de porqué las jirafas tienen el cuello largo, si sus ancestros tenían cuello corto. La explicación Lamarckiana a este fenómeno, es que en algún momento del pasado las condiciones del medio ambiente cambiaron, de tal modo que arbustos y pastos de los que se alimentaban las jirafas de cuello corto, escasearon, lo que llevó a las jirafas de cuello corto a explorar nuevas fuentes de alimento como lo son árboles de mayor altura.
El esfuerzo constante por alcanzar alimento que se encontraba a una altura mayor, provocó que el cuello de las jirafas paulatinamente se alargara, y de esta manera se dio origen al cuello tan largo de las jirafas que conocemos en la actualidad.
¿Cuál es la idea que prevalece actualmente?
Charles Darwin plasmó su teoría de la evolución biológica con la publicación del libro “El origen de las especies”, en el que propuso como motor de la evolución a la selección natural a través de la diversidad de especies, de esta manera se pudo dar una explicación más aceptable al porqué las jirafas tienen el cuello largo.
Se planteó un escenario donde existían tanto jirafas de cuello corto como jirafas de cuello largo, de igual manera ocurrió un cambio en el medio ambiente que provocó la escasez de pastos y arbustos, dejando como único alimento disponible los árboles. Encontrándose la única fuente de alimento disponible a altura mayor, sólo las jirafas de cuello largo pudieron alimentarse y sobrevivir, y las jirafas de cuello corto no pudieron alimentarse, lo que llevó a su desaparición.
La supervivencia de las jirafas de cuello largo se debió a que presentaron un carácter (cuello largo) que en ese momento representó una ventaja adaptativa, que tuvo como consecuencia la preservación de su especie. Y fue así que la idea Darwiniana de la evolución ha prevalecido hasta nuestros días, permitiéndonos explicar cómo es que la vida logró transformarse a como la conocemos en la actualidad.
Genética y epigenética
Todos al menos una vez, hemos escuchado hablar sobre los genes y de cómo las características de padres a hijos son transmitidas por éstos. Darwin tenía idea de cómo las especies evolucionaban, pero desconocía como estas características que permitían la supervivencia de determinada especie era heredada de una generación a otra. Hasta que fue descubierto el trabajo de Gregor Mendel, quien, analizando de qué manera y cómo se transmitían características específicas (como el color de las flores) de una generación a otra de plantas de chícharo, pudo identificar a los genes a los que él denominó “factores” como los responsables de la herencia (no fue hasta 1909 que el botánico Wilhem Johannsen acuñó el término “gen” a la unidad física y funcional de la herencia). Así nace la genética como el área de la ciencia que estudia a los genes, su variación y su heredabilidad en los organismos vivos.
En la actualidad, resumiendo la teoría Darwiniana y Mendeliana, sabemos que el motor de la evolución es la mutación en los genes.
Los genes son secuencias de ADN, que a su vez está formado por una combinación de 4 bases nitrogenadas (G= guanina, C= citosina, A= adenina y T= timina), además los genes tienen secuencias de bases nitrogenadas de tamaño variable (por ejemplo, el gen CHS mide 1453 pares de bases, el gen SOD mide 788 pares de bases y el gen PR1 mide 902 pares de bases, todos genes, se miden en pares de bases ya que el ADN es una cadena doble). Haciendo analogía con nuestro lenguaje, los genes serían palabras escritas con un alfabeto que en este caso tendría sólo 4 letras G, C, T y A. De acuerdo con la secuencia del gen (el orden de las bases) será la función específica que éste tendrá. Mutaciones (cambios) en la secuencia de los genes, generan versiones diferentes del mismo gen, a lo que llamamos alelos. Un ejemplo de mutación puede ejemplificarse de la siguiente manera:
Gen original: ATGTACTAGCATATCT
Gen mutado: ATGTTCTAGCATATCT
Entonces tenemos 2 versiones del gen, el gen original y el gen mutado, la diferencia entre ambos es el cambio en una base (la quinta base del gen original es A y en el gen mutado es T).
Suponiendo que el gen original da lugar a la pigmentación de un animal, la mutación de una base (marcada en rojo) podría ocasionar que el animal pierda la pigmentación. De esta manera un gen puede tener dos o más versiones o alelos que se heredan de una generación a otra, generando la diversidad de organismos vivos que conocemos en la actualidad.
La secuencia de los genes no es la única información heredable en los organismos vivos. Señales del medio ambiente pueden generar modificaciones sobre los genes, que no modifican la secuencia (como lo hacen las mutaciones), sólo su manifestación, mediante un “marcaje químico”, a este marcaje de genes se le conoce como regulación epigenética.
La epigenética es esencialmente información adicional colocada encima de las letras (A, C, G, y T) que conforman el ADN. Sí se considera a la secuencia de ADN como un texto instructivo que explica cómo construir a un ser vivo, la epigenética sería como sí se tomaran marcatextos de diferentes colores para marcar el texto de diferentes maneras, para recalcar la importancia de las diferentes partes del texto. Por ejemplo, alguien podría usar color azul para resaltar partes del texto que necesitan ser leídas con mayor cuidado y color rojo para marcar partes que no son tan importantes.
En este “marcaje de ADN”, las marcas son agregadas al ADN por proteínas particulares en las citosinas (las letras C) de genes específicos, generando versiones diferentes de genes, denominados epialelos. Por ejemplo, el ADN puede ser marcado con unas moléculas pequeñitas llamadas grupos metilos en las citosinas. Existen proteínas que buscan y se unen a estas áreas metiladas, apagando los genes que se encuentran en estas áreas, así entonces éstas metilaciones son como las partes del texto marcadas con el marcatextos rojo, diciéndole a la célula “no necesitas a éstos genes ahora”.
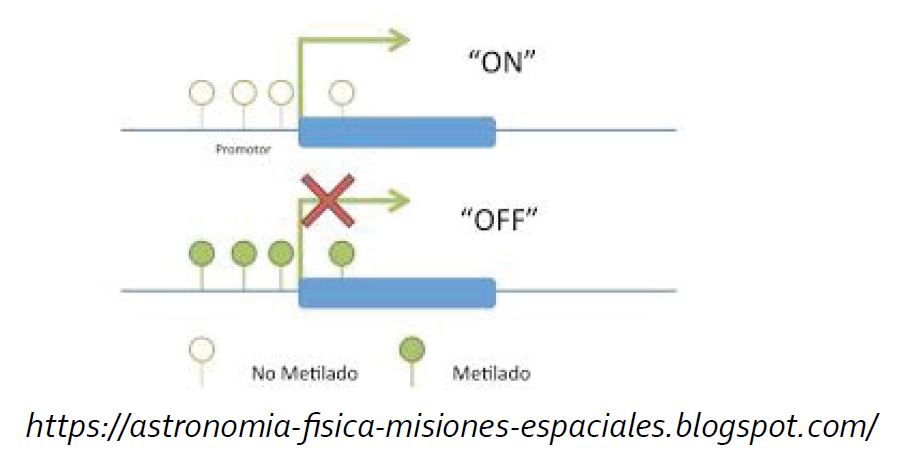
El ADN es una molécula de gran tamaño que necesita ser empaquetada para poder caber dentro de la célula. Existen proteínas llamadas histonas que sirven a manera de carrete para que el ADN se enrolle alrededor, formando una estructura llamada nucleosoma.

Grupos metilo y otras moléculas pequeñas pueden ser pegadas sobre diferentes zonas en las proteínas histonas, teniendo un efecto diferente. Algunas marcas o etiquetas en algunas zonas aflojan la unión entre el ADN y la histona, haciendo al ADN más accesible para que las proteínas responsables de la expresión de los genes puedan unirse y así encender los genes en ésta región, esto sería como la parte del texto marcada con marcatextos azul diciendo a la célula “éstos genes son necesarios ahora”.
¿Qué son los epilalelos?
Los epialelos son iguales en la secuencia de bases a los alelos de los genes originales, la diferencia entre ellos es el marcaje o etiquetado (como las metilaciones) lo que provoca que se manifiesten de manera distinta, generando variantes de los genes que no dependen de la secuencia de bases, por lo tanto tampoco dependen de la mutación. Un ejemplo de la variación producida por un cambio epigenético puede observarse en la planta Linaria vulgaris, la cual cambia la simetría de sus flores por el marcaje en un gen particular, el gen Lcyc .
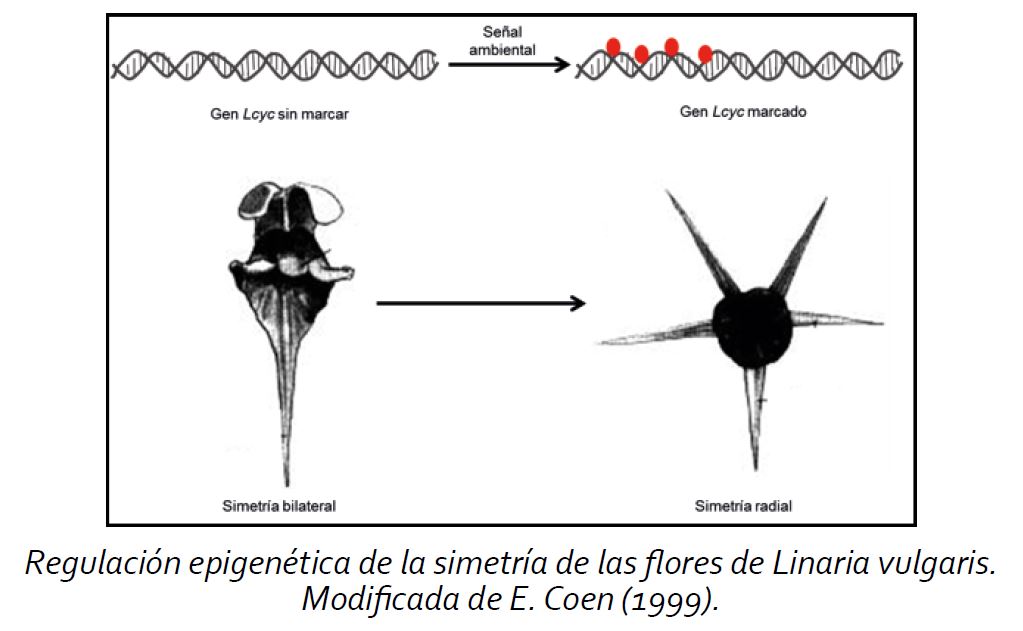
Las plantas que no poseen el marcaje del gen Lcyc (estado natural) tienen una simetría bilateral y aquellas plantas que poseen el marcaje del gen presentan una simetría radial; ambas variantes de las plantas poseen la misma secuencia del gen, la única diferencia que presentan es el marcaje de el gen Lcyc.
Algunos epialelos son transmitidos fielmente. Por lo tanto, en los casos donde los epialelos tienen consecuencias favorables en la adaptación para el organismo, pueden representar una forma adaptativa heredable conocida como herencia Lamarckiana (hipótesis de que un organismo puede pasar rasgos adquiridos durante su ciclo de vida a su progenie). Esto puede ofrecer un incremento en la supervivencia de subsecuentes generaciones en condiciones de estrés medioambientales similares. Usando el ejemplo de Linaria vulgaris, sí el cambio en la simetría de la flor representara una ventaja que le favoreciera (por ejemplo, en la reproducción), la variante preservaría sus poblaciones, asegurando su supervivencia.
De esta manera, los epialelos se suman a las mutaciones como fuente de variación de los organismos, así como una fuerza motora adicional de la evolución.
Masuelli R. y Marfi C. (2011). Variabilidad epigenética en plantas y evolución. J Basic Appl Genet 22:1852-6233.
http://www.scielo.org.ar/scielo.php?script=sci_arttext&pid=S1852-62332011000100001.
Gutiérrez-Velázquez, M.V. (2015). Metilación del ADN vegetal, principal mecanismo epigenético en plantas. Vidsupra visión científica 7:1-5. http://www.ciidirdurango.ipn.mx/REVISTA_VIDSUPRA/Documents/tabla%20contenido%20v7%20n1/ART1_METILACION.pdf
Pikaard, C. y Scheid O. (2017). Epigenetic regulation in plants. Cold Spring Harb Perspect Biol 6:a0199315.
http://cshperspectives.cshlp.org/content/6/12/a019315.full.pdf+html
El M.C. Christian Hernández Soberano es estudiante del Programa Institucional del Doctorado en Ciencias Biológicas en la Opción de Biología Experimental.
Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.
D.C. Eduardo Valencia Cantero es profesor e investigador, ambos del Instituto de investigaciones Químico Biológicas de la Universidad Michoacana de San Nicolás de Hidalgo.